
我々が不均衡進化理論を1992年に発表してから既に30年以上が経ちました。しかし、進化研究者の間では未だに定説とはなっていません。投稿論文の査読者から、「古澤らの説は数理的にどこが間違っているとは指摘できないが、彼らの結論は“遺伝学的熱力学”の基本法則に悖るので受理できない。」という返事が来たり、「この論文の著者らは長年同じ研究を続けているが、世界の研究者の中で彼らの研究を信じている者は誰もいない。」という、何とも取り付く島もない反応もあります。今もこの状況は大して変わりません。最近は、明らかに生成AIを使ったと思われるコメントを送ってくる査読者も散見されます。
一方、不均衡進化理論は発表当初から物理学者の注目を引きました。最初の論文発表と同じ年に、当時ゲッチンゲン市のマックス・プランク生物物理・化学研究所の所長であったM. アイゲン博士(1967年度ノーベル化学賞受賞)のお招きで特別講演の機会を得ました。教授は遺伝や進化にも興味があり、準種(Quasi-species)の提唱者としても有名です。講演後、博士の御希望で共同研究者の土居洋文博士(当時、富士通)共々、一日滞在を延長して議論を続けました。また、アイゲン博士には米国科学アカデミー紀要へのDNA型遺伝アルゴリズムに関する論文の推薦の労を執っていただきました(文献1)。上記の講演会で話した内容は、2003年のPhysical Review E誌(非線形物理・生物物理分野)にアイゲン・シュスターの準種を発展させた不均衡変異の数理的解析の結果として掲載されています(文献2)。2003年11月には、京大基礎短期研究会主催の講演会『量子力学とカオス:基礎問題からナノサイエンスまで』に招かれ講演をしました。英語とその邦訳を交互に混ぜ合わせて話しましたが、会場や座長からの質問の内容が理解できず、もどかしい時間を過ごした記憶があり、自身の不勉強を自覚しました。
生命科学分野からの反応が全くなかったわけではありません。英国の世界初の理論生物学雑誌であるJournal of Theoretical Biology誌 の編集長であったL. ウオルパート博士(組織分化の“フランス3色旗説”で有名)から、『査読者は全員否定的だが、自分は受理したいので図を一つ外してくれないか?』という内容の直接電話があり、一も二もなく受諾しました。こうして投稿を始めてから凡そ3年を経過した後に、不均衡進化理論の最初の論文が日の目を見ることになったのです。それまでにNature誌、Science誌を始めいくつかの有名誌に投稿しましたが、全て門前払いでした。もう一つエピソードを。確か前にもコラムに書いたと思うのですが、最初の論文が出た頃、ある新聞社の若い記者の方と立ち話をしているとき、『この理論は“元本保証の多様性拡大”だな。』と呟かれました。その言葉に感銘を受けて、それ以来このフレーズを不均衡進化理論のキャッチコピーとして使わせていただいております。
でも、どうしてこれほど分野によって極端に評価が分かれるのでしょう?私事で恐縮ですが、わが家には妻と娘が一人います。ある日の夕食後、二人に不均衡進化理論の話をごく分かりやすく説明する機会がありました。理系の妻の反応は「少しも理解できない、すぐにメンデルの遺伝の法則が出てきて頭が混乱する。」という返事でした。ところが文系の娘からは、「なるほど、よくわかった。」と言う返事をもらいました。このエピソードからの判断ですが、我々の進化理論を理解するには、学校で習う生物学の知識がかえって邪魔をすることがよく分かりました。そう言えばほかの分野でも、専門知識がかえって邪魔をするというフレーズはよく耳にします。
生命科学者、特に遺伝学者や進化の研究者は、突然変異という言葉を聞くと、メンデルの遺伝の法則→遺伝子浮動→中立進化→自然選択→進化、と反射的に思考が一挙に進んでしまうのでしょう。単なる筆者の想像にすぎませんが、このロジックは進化論の本流として確固たる地位を占めていますから、今更他のファクターを考慮する必要性は毛頭ないと感じてしまうのではないでしょうか?ここで読者に知っていただきたいことは、メンデル遺伝には新しく入る変異は全く考慮されていない点です。
これ迄の筆者のコラムを継続してお読みいただいている読者の皆様には意外に思われるかもしれませんが、不均衡進化理論はネオ・ダーウィニズムとは敵対するものではありません。その主な理由は、近代のネオ・ダーウィニズムは単なる理屈(数理理論)に留まるのではなく、その理論の大部分が生体材料の分析や観察の結果をもとに確実に裏付けされているからです。ただ変異の捉え方が両者で違うだけです。しかし、以下に述べますように、この立場の違いこそが進化の解釈に決定的な影響を及ぼすことになります。
本コラムのシリーズで繰り返し説明してきましたように、ネオ・ダーウィニズムも不均衡進化理論も平均変異率(トータルの変異率μ)は同じです。ただし、ネオ・ダーウィニズムでは変異はランダムに平均して入るものと仮定しています。直感でもすぐに分かるように、平均変異数がゲノム当たり1個を超すとエラー・カタストロフィーを起こして集団は自滅してしまうという数理上の足枷(変異の閾値)が現れます。何故なら、変異の中には必ず致死や有害変異が混ざっているからです。従って、進化は極めてゆっくりと進まざるを得なくなり、とても進化のジャンプを説明することは出来ません。しかし事実として、カンブリア爆発のような急速進化の証拠があります。また、ヒトの赤ちゃんは両親から約70個の変異を受け継ぎます。計算上のことですが、ヒトは“2度死ぬ”ことになります(文献3)。逆に、進化が停止したように見える生ける化石の存在理由もうまく説明できていません。このように、ネオ・ダーウィニズムには大きな矛盾が見られます。この難題解決の方法はいろいろ考えられてはいますが、納得できる説明はありません。この難題を解くためには、何としても、集団の自滅を回避して変異率を上げ、同時に変異率が低い個体も担保できる隠されたトリックを探さねばなりません。
筆者がDNA複製の分子モデルを見て閃いたのは、DNAの複製で生じる2匹の娘DNA間の変異率の差に注目すればこのトリックが明らかになるかもしれないという事でした。この変率の差をFidelity difference(FD:忠誠度の差)と表現します。FDはこれまで誰も注意を払わなかった突然変異率に関する新しい“変数”です。ネオ・ダーウィニズムの概念にFDを加えるという極めて簡単な操作で、問題は見事に解決されました。その効果は以下の3つに要約することが出来ます。
1)適切な条件下では、変異の閾値の上昇或いは解消が起こり、平均変異率μがどれほど上昇しても、変異の入らない個体と過剰の変異が入った個体を同時に創り出すことができる(元本保証の多様性拡大)。
2)一度集団内に現れた遺伝子型の確実な継承。
3)集団内に蓄積された有害突然変異の効率的な除去。
これらの3つの効果が顕著に現れる条件は、2匹の娘DNAのどちらか一方の変異率が有意に低い時です。図を参照ください。
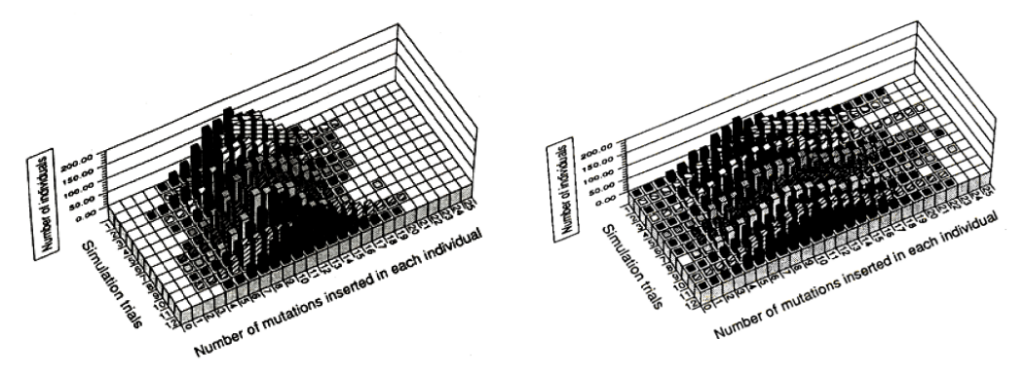
左の図に示すように、1個体から始まる集団の成長を追っていくと、従来の均衡変異の下では変異体の分布はベル状になり、変異0の元の個体は存在しません。世代が進んで行くと、ベル状の山は大きくなり、その形状を保ちながら右へ右へと移動します。一方、不均衡変異の場合には、何時までたっても元の変異0の個体が存在し続けるだけではなく、より多くの変異が蓄積した個体も出てきます。結果として、山は平べったく裾野の広い緩やかな丘陵のような形になって行きます(右図)。このように、カンブリア爆発のような急速進化や、シーラカンスやカブトガニのように進化が止まったように見える生ける化石の存在など、ダーウィン進化で指摘されている諸々の矛盾の大部分が説明可能となります(文献3)。
生物集団をヒトの赤ちゃんに、不均衡変異の創り出す場(筆者らは最近この“場”を“不均衡準種;Disparity quasi-species”と名付けています)を“揺りかご”に喩えることができるでしょう。ネオ・ダーウィニズムでは、赤ちゃんは均衡変異という固定ベッドの上で育てられていました。しかし不均衡準種の場では、“揺りかご”の中で大切に育てられています。お母さんが揺り動かす心地よい振動の中で、赤ちゃんはきっとより健やかに育つでしょう。実際の揺りかごと異なる点は、この“揺りかご”は赤ちゃん自ら作り出した物で、赤ちゃん自身が揺すっているのです。この力がFDという訳です。このように、集団の背後では途切れることのない変異の波が押し寄せています。この波の原因となっているFDが確かに存在する実例が、かなりの数報告されています。以前、我々が大腸菌で調べた結果によりますと、FDの値は11倍でした(文献4)。
幸運にも我々は、“変異の振動波”の進化促進効果の一端を垣間見ることが出来ました。従来の考えではその名が示すように、突然変異はどこからか突然やって来るものと認識されていました。一方、不均衡変異理論では、変異を起こしやすいポテンシャルの高い“場”と、そうでない低い“場”があり、それらがDNA複製機構と強くカップリングしているとするのが我々の主張です。ここまで筆を進めてきて、これは人を説得するのは大変だなとの想いがますます募ってきました。しかし、FDの存在と進化の関係の解明は道半ばです。理解していただく努力を続けながら研究を進めていくしかありません。現在、複製に伴ってFDが現れる“場”の実体を明らかにしようと研究を続けています。
著者の独り言。『そうか、物理学者や物理化学者は“場”の概念には慣れている。しかし、生物学者には「同じ遺伝情報を持つDNA鎖であっても、複製に伴って変異が起こり易いポテンシャルを持ったものと、そうでないものが現れる。」と言う“場”の概念が入ったストーリーは理解し難いのかな?』。
2026年3月19日
古澤 満
文献:
- Wada, K., Doi, H., Tanaka, S., Wada, Y., & Furusawa, M. (1993).
A neo-Darwinian algorithm: Asymmetrical mutations due to semiconservative DNA-type replication promote evolution.
Proceedings of the National Academy of Sciences, 90(24), 11934–11938
- Aoki, K., & Furusawa, M. (2003).
Increase in error threshold for quasispecies by heterogeneous replication accuracy. Physical Review E, 68(3), 031904
- 不均衡進化理論とネオ・ダーウィニズム[第57回 古澤満コラム]
- Iwaki, T., Kawamura, A., Ishino, Y., Kohno, K., Kano, Y., Goshima, N., Yara, M., Furusawa, M., Doi, H., & Imamoto, F. (1996).
Preferential replication-dependent mutagenesis in the lagging DNA strand in Escherichia coli.
Molecular and General Genetics MGG, 251(6), 657-664.